


Abstract:
A Japan-based research team led by Kanazawa University has identified the MAPK Erk5 as a novel player controlling skeletogenesis. Their research sheds light on the question of how the complex our skeleton generates.
[Background]
There are more than 200 bones in the human body. Bone is formed during embryonic and postnatal skeletogenesis by two distinct, well-organized processes, intramembranous and endochondral ossification. Mesenchymal stem cells differentiate into chondrocytes to form a cartilaginous template, which, for long bones, induces bone formation through endochondral ossification. Extracellular signal-regulated kinase 5 (Erk5), which is a member of the mitogen-activated protein kinase (MAPK) family, is phosphorylated by MAPK/Erk kinase-5 (Mek5) to regulate the function of various cell types. Although the MAPK Erk1/2 pathway is well known for regulating skeletogenesis, the in vivo physiological role of the Mek5/Erk5 pathway in skeletal development has been largely unclear to date, because of the early embryonic lethality of global Erk5 knockout mice.
[Results]
By using cell-specific mouse genetics approaches, Takashi Iezaki, Katsuyuki Kaneda and Eiichi Hinoi at Kanazawa University, Japan, and colleagues revealed that Erk5 plays a crucial role in skeletogenesis in vivo. Paired-related homeobox 1 (Prx1) is expressed in mesenchymal stem cells in the limb buds. Mesenchymal stem cell-specific Erk5 knockout embryos (Prx1-Cre;Erk5fl/fl embryos) displayed wider long bones and impaired mineralization of the metatarsal. In contrast, chondrocyte-specific Erk5 knockout embryos (Col2a1-Cre;Erk5fl/fl embryos) recapitulated only the wider long bone phenotype of Prx1-Cre;Erk5fl/fl embryos. Accordingly, the investigators revealed that Erk5 controls 1) early chondrogenic differentiation of mesenchymal stem cells, including mesenchymal condensation, through its expression in skeletogenic progenitors, and 2) chondrogenic differentiation after the formation of mesenchymal condensations through its expression in chondrocytes.
Subsequent biochemical analyses found that Erk5 directly phosphorylates Smad1 at Ser206 in the linker region, which is known to trigger its proteasomal degradation in a Smad-specific E3 ubiquitin ligase 1 (Smurf1)-dependent manner. In addition, Erk5 directly phosphorylates Smurf2 at Thr249, leading to accelerated proteasomal degradation of Smad proteins (Smad1, 2 and 3) (Figure 1). The study demonstrated that Smads transcriptionally activate the expression in mesenchymal stem cells of sex-determining region within the Y-type high-mobility group box protein 9 (Sox9), which is the principal transcription factor involved in skeletogenesis. Moreover, using mouse genetic rescue experiments, the investigators revealed that Sox9 is a critical mediator of Erk5-dependent skeletogenesis.
[Significance and future prospects]
In conclusion, the Mek5/Erk5 pathway is critical for skeletogenesis in vivo through its expression in mesenchymal stem cells and modulation of Smad protein stability (Smad1, 2, and 3) via Smurf activity (Smurf1 and 2) (Figure 2). These findings improve our understanding of the molecular mechanisms underlying skeletal development and spur the development of drugs targeting human cartilage diseases associated with abnormal chondrocyte differentiation and maturation. Moreover, since the Smurf2/Smads cascade is associated with other diseases, including cancer and ageing, the newly identified Mek5/Erk5/Smurf2/Smads/Sox9 cascade is a candidate target for developing drugs to treat a variety of human diseases.
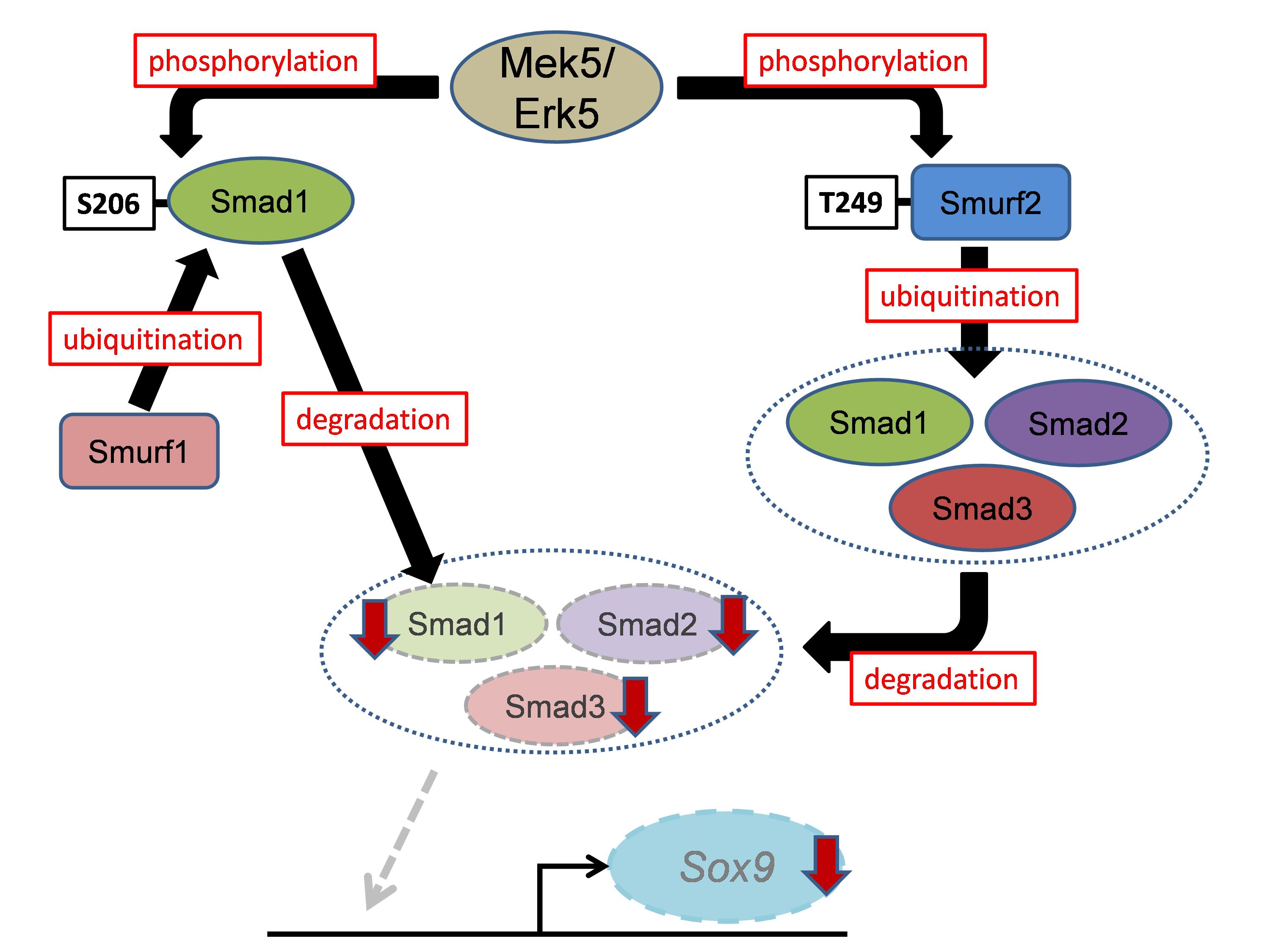
Figure 1. Schematic model of this study.
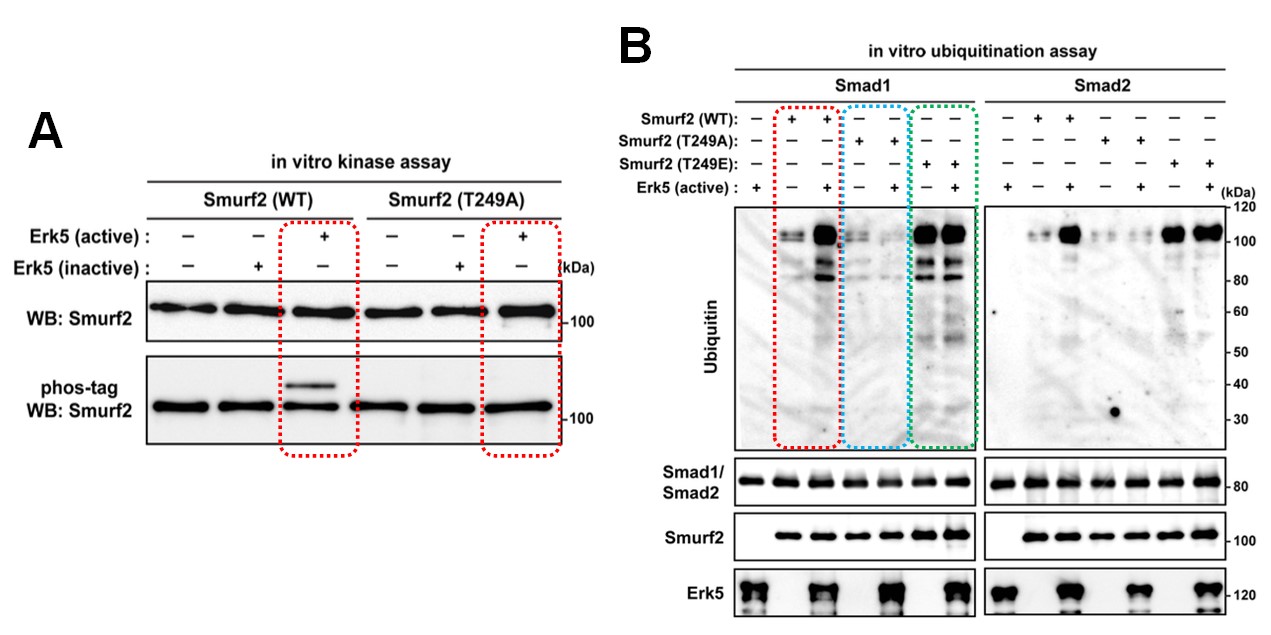
Figure 2.
(A) In vitro kinase assay. Recombinant Smuf2 proteins were incubated with active or inactive Erk5, and subsequent Phos-tag SDS-PAGE, followed by immunoblotting. Recombinant Smurf2(WT) was phosphorylated by Erk5(active) but not by Erk5(inactive), whereas Smurf2 phosphorylation by Erk5 was stopped completely by Smurf2(T249A), in which the threonine was replaced with an alanine. (B) In vitro ubiquitination assay. Recombinant Smuf2 proteins and Smad proteins were incubated with active Erk5 in the presence of E1 and UbcH5c, followed by SDS-PAGE. By incubating Smurf2(WT) with ubiquitin, E1 and UbcH5c caused Smad protein ubiquitination in the presence of Erk5 despite the weak ubiquitination signal noted in the absence of Erk5. The reaction with Smurf2(T249A) produced a faint ubiquitination signal in the presence of Smads irrespective of the presence of Erk5, whereas extensive ubiquitination was observed when Smads were incubated with Smurf2(T249E), in which threonine was replaced with glutamate, in the absence of Erk5.
Article
The MAPK Erk5 is necessary for proper skeletogenesis involving a smurf-smad-Sox9 molecular axis
Journal: Development
Authors: Takashi Iezaki, Kazuya Fukasawa, Tetsuhiro Horie, Gyujin Park, Samuel Robinson, Michio Nakaya, Hiroyuki Fujita, Yuki Onishi, Kakeru Ozaki, Takashi Kanayama, Manami Hiraiwa, Yuka Kitaguchi, Katsuyuki Kaneda, Yukio Yoneda, Takeshi Takarada, X. Edward Guo, Hitoshi Kurose, Eiichi Hinoi
DOI: 10.1242/dev.164004
Funders
This work was supported in part by the Japan Society for the Promotion of Science (16H05131, 17KT0051 and 18H04971 to E.H.); and the Japan Agency for Medical Research and Development (17824969 to E.H.).